



Одним из самых известных и наиболее изученных из палеонтологических доказательств эволюции является филогенетический ряд современных однопалых копытных. Множественные палеонтологические находки и выявленные переходные формы создают научную доказательную базу данного ряда. Описанный русским биологом Владимиром Онуфриевичем Ковалевским еще в 1873 году филогенетический ряд лошади и сегодня остается «иконой» эволюционной палеонтологии.
Эволюция через века
В эволюции филогенетические ряды — это последовательно сменявшие друг друга переходные формы, приведшие к формированию современных видов. По количеству звеньев ряд может быть полным или частичным, однако наличие последовательных переходных форм является обязательным условием их описания.
Филогенетический ряд лошади относят к доказательствам эволюции именно благодаря наличию таких последовательных форм, сменяющих друг друга. Множественность палеонтологических находок наделяет его высокой степенью достоверности.
Доказательства эволюции
Ч. Дарвин доказал историческое развитие живой природы, но в последующие годы сбор прямых и косвенных доказательств эволюции продолжался.
Филогенетические ряды
Выдающиеся достижения в накоплении прямых доказательств эволюции принадлежат отечественным ученым, прежде всего В. О. Ковалевскому. Работы В. О. Ковалевского были первыми палеонтологическими
исследованиями, которыми удалось показать, что одни виды происходят от других.
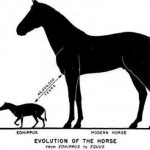
Исследуя историю развития лошадей, В. О. Ковалевский показал, что современные однопалые животные происходят от мелких пятипалых всеядных предков, живших 60—70 млн. лет назад в лесах. Изменение климата Земли, повлекшее за собой сокращение площадей лесов и увеличение размеров степей, привело к тому, что предки современных лошадей начали осваивать новую среду обитания — степи. Необходимость защиты от хищников и передвижений на большие расстояния в поисках хороших пастбищ привела к преобразованию конечностей — уменьшению числа фаланг вплоть до одной (рис. 14). Параллельно изменению конечностей происходило преобразование всего организма: увеличение размеров тела, изменение формы черепа и усложнение строения зубов, возникновение свойственного травоядным млекопитающим пищеварительного тракта и многое другое.
Рис. 14. Преобразование конечностей предков лошадей из пятипалой в трехпалую и затем в однопалую. Последняя конечность — современная лошадь.
В. О. Ковалевский обнаружил последовательные ряды ископаемых форм лошадиных, эволюция которых совершалась в указанных направлениях. Такие ряды видов, последовательно сменяющих друг друга, называются филогенетическими
 Читайте также:
Читайте также:

и свидетельствуют о существовании эволюционного процесса.
Переходные формы
Установление факта постепенного эволюционного развития в линиях невысокого систематического ранга (ряды ископаемых лошадей, слонов, моллюсков) показало существование преемственности между современными и ископаемыми видами. Не в силах опровергнуть эти факты, противники эволюционной теории утверждали, что систематические группы более высокого ранга не могли произойти друг от друга, а явились результатом отдельного акта творения. Поэтому особый интерес представляют ископаемые формы, сочетающие признаки древних и более молодых групп высокого систематического ранга. Такие формы называются переходными.
Примером их могут служить кистеперые рыбы, связывающие рыб с вышедшими на сушу четвероногими земноводными; семенные папоротники — переходная группа между папоротиикообразными и голосеменными и др.
Существование переходных форм между разными типами, классами, отрядами показывает, что постепенный характер исторического развития свойствен не только низшим, но и высшим систематическим категориям.
Гомологнчные и аналогичные органы, рудименты и атавизмы
Сравнительно-анатомические исследования показали, что строение передних конечностей некоторых позвоночных, например ласты кита, лапы крота, крокодила, крылья птицы, летучей мыши, руки человека, несмотря на выполнение совершенно разных функций, в принципиальных чертах строения сходны. Некоторые кости в скелете конечностей могут отсутствовать, другие срастаться, относительные размеры костей могут меняться, но их гомология,
т. е. сходство, основанное на общности происхождения, совершенно очевидна. Гомологичными называются такие органы, которые развиваются из одинаковых эмбриональных зачатков сходным образом (рис. 15).
Наличие у организмов разных групп (классов, семейств и т. д.) гомологичных органов дает возможность установить степень родства между ними, проследить их эволюцию. Видоизменение органов, имеющих общее происхождение, объясняется дивергенцией по признаку строения данного органа в связи с приспособлением к среде обитания.
-
Читайте также:

Не всякое сходство органов свидетельствует в пользу их родства. Крыло бабочки и крыло птицы выполняют сходную функцию, но их строение совершенно различно. Сходство вызвано образом жизни, приспособлением к полету, возникшим независимо друг от друга у бабочек и птиц, а не родственным происхождением этих форм. Органы, имеющие внешнее сходство, вызванное сходными приспособлениями к сходным условиям жизни, но различное строение, называются аналогичными. Аналогичные органы
возник-
Рис. 15. Сходство строения передних конечностей обезьяны, летучей мыши, тюленя и лошади свидетельствует об их происхождении от единой предковой формы или в результате конвергенции
— схождения признаков и не свидетельствуют о родстве между организмами.
Рис. 16. Конвергенция по форме тела у прыгающих млекопитающих.
Некоторые органы или их части не функционируют у взрослых животных и являются для них лишними — это рудиментарные органы,
или рудименты. Наличие рудиментов, так же как и гомо-логичных органов, свидетельствует об общности происхождения живых форм. Задние конечности у кита, скрытые внутри тела,— рудимент, доказывающий наземное происхождение его предков. У человека тоже известны рудиментарные органы: мышцы, двигающие ушную раковину, рудимент третьего века и т. п.
У некоторых организмов рудиментарные органы могут развиться до органов нормальных размеров. Такой возврат к строению органа предковых форм называется атавизмом. Среди тысяч однопалых лошадей изредка попадаются особи, у которых развиты маленькие копытца II и IV пальцев. Известны случаи появления атавистических признаков и у человека: рождение детей с первичным волосяным покровом, с длинным хвостиком и т. д. Возникновение атавизмов указывает на возможное строение того или иного органа у предковых форм.
Сходство зародышевого развития позвоночных

Факт единства происхождения живых организмов был установлен на основе эмбриологических исследований. Все многоклеточные животные развиваются из одной оплодотворенной яйцеклетки. В процессе индивидуального развития они проходят стадии дробления, образования двух- и трехслойного зародышей, формирования органов из зародышевых листков. Сходство зародышевого развития животных свидетельствует о единстве их происхождения.
С особой отчетливостью сходство эмбриональных стадий выступает в пределах отдельных типов и классов. Так, на ранних стадиях развития у зародышей позвоночных (рыбы, ящерицы, кролика, человека) наблюдается поразительное сходство: все они имеют головной, туловищный и хвостовой отделы, зачатки конечностей, по бокам тела — зачатки жабр (рис. 17).
По мере развития зародышей черты различия выступают все более явственно. Причем вначале проявляются признаки класса, к которому относятся зародыши, затем признаки отряда и на еще более поздних стадиях — признаки рода и вида. Эта закономерность в развитии зародышей указывает на их родство, происхождение от одного ствола, который в ходе эволюции распался на множество ветвей.
Биогенетический закон
Основываясь на приведенных выше, а также множестве других фактов, немецкие ученые Ф. Мюллер и Э. Геккель во второй половине XIX в. установили закон соотношения онтогенеза, который получил название биогенетического закона. Согласно этому закону каждая особь в индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез), или, короче, онтогенез есть краткое повторение филогенеза.
Однако за короткий период индивидуального развития особь не может повторить все этапы эволюции, которая совершалась тысячи или миллионы лет. Поэтому повторение стадий исторического развития вида в зародышевом развитии происходит в сжатой форме, с выпадением ряда этапов.
Рис. 17. Сходство начальных стадий онтогенеза позвоночных свидетельствует о родстве и указывает на пройденные в процессе филогенеза этапы
Кроме того, эмбрионы имеют сходство не со взрослыми формами предков, а с их зародышами. Так, в онтогенезе млекопитающих и рыб имеется этап, на котором у зародышей образуются жаберные дуги. У зародыша рыбы на основании этих дуг образуется орган дыхания — жаберный аппарат. В онтогенезе млекопитающих повторяется не строение жаберного аппарата взрослых рыб, а строение закладок жаберного аппарата зародыша, на основе которых у млекопитающих развиваются совершенно иные органы (хрящи гортани и трахеи). В разработке теории онтогенеза выдающуюся роль сыграли исследования академика А. Н. Северцова. Он доказал, что изменения исторического развития обусловлены изменениями хода зародышевого развития. Наследственные изменения затрагивают все стадии жизненного цикла, в том числе и зародышевый период. Мутации, возникающие в ходе развития зародыша, как правило, нарушают взаимодействие в организме и ведут к его гибели. Однако мелкие мутации могут оказаться полезными и тогда сохранятся естественным отбором. Они передадутся потомству, включатся в историческое развитие, влияя на его ход.
Примеры филогенетических рядов
Ряд лошадей не единственный среди описанных примеров. Хорошо изучен и имеет высокую степень достоверности филогенетический ряд китов и птиц. А спорным в научных кругах и наиболее используемым при различных популистских инсинуациях является филогенетический ряд современных шимпанзе и человека. Споры по поводу недостающих здесь промежуточных звеньев не утихают в научной среде. Но сколько бы ни было точек зрения, бесспорным остается значение филогенетических рядов как доказательств эволюционной приспособляемости организмов к изменяющимся условиям окружающей среды.
-
Читайте также:

Корреляция эволюции с окружающей средой
Скелет и внешнее строение предков современных форм лошадей и слонов демонстрируют четкую взаимосвязь с изменениями температурного режима, а значит, и ландшафта нашей планеты. Изменения климата уменьшали лесные массивы, увеличивая площадь степей и саванн. Предкам лошадей и слонов приходилось приспосабливаться, что еще раз доказывает значение филогенетических рядов в эволюционной приспосабливаемости организмов к изменяющимся условиям окружающей среды. Менялись строение скелета, строение зубной системы и размеры животных.
Промежуточные звенья
Порядка 25 миллионов лет назад климат на планете изменился, и на смену лесам стали приходить степные просторы. В миоцене (20 миллионов лет назад) появляются мезогиппус и парагиппус, уже более похожие на современных лошадей. А первым травоядным предком в филогенетическом ряду лошади принято считать мерикгиппуса и плиогиппуса, которые выходят на арену жизни 2 миллиона лет назад. Гиппарион – последнее трехпалое звено
Этот предок жил в миоцене и плиоцене на равнинах Северной Америки, Азии и Африки. Эта трехпалая лошадка, напоминающая газель, еще не имела копыт, но могла быстро бегать, ела траву и именно она заняла огромные территории.
Чем питаются?
Обзор филогенеза позволяет узнать все о лошадях. Эволюция предшествующих видов имела четкое направление. Все они развивались в условиях отступления лесов и наступления степей и лесостепей. А это означает, что предшествующим видам приходилось эволюционировать не только в сторону увеличения размеров тела. Они должны были менять и свой рацион.
Современные лошади и их ближайшие предки уже не могли питаться сочными плодами. Им пришлось перейти на траву и зерна злаков, поэтому домашние кони питаются пшеницей, овсом, ячменем и другими злаками, хотя не отказываются при возможности от яблок, моркови, огурцов, тыквы и других сочных плодов.
Знакомая нам лошадь
Современная лошадь как конечный этап филогенетического ряда появилась в конце неогена, а в конце последнего ледникового периода (около 10 тысяч лет назад) в Европе и Азии уже паслись миллионы диких лошадей. Хотя усилия первобытных охотников и сокращение пастбищ сделали дикую лошадь редкостью уже 4 тысячи лет назад. Но два ее подвида – тарпан в России и лошадь Пржевальского в Монголии – сумели продержаться гораздо дольше всех остальных.
Биогеографические доказательства эволюции
Сравнение фаун и флор разных континентов. Яркое свидетельство эволюционного процесса — распространение животных и растений по поверхности нашей планеты. Во все времена путешественников и натуралистов поражало своеобразие растительного и животного мира Австралии, Южной Америки, океанических островов и вызывало интерес сходство фауны некоторых материков, например Северной Америки и Евразии. А. Уоллес привел все сведения в систему и выделил шесть зоогеографических областей:
1) Палеоарктическую, охватывающую Европу, Северную Африку, Северную и Среднюю Азию, Японию;
2) Неоарктическую, включающую Северную Америку;
3) Эфиопскую, включающую Африку к югу от пустыни Сахара;
4) Индомалайскую, охватывающую Южную Азию и Малайский архипелаг;
5) Неотропическую, занимающую Южную и Центральную Америку;
6) Австралийскую, включающую Австралию, Новую Гвинею, Новую Зеландию, Тасманию, Соломоновы острова и Новую Каледонию.
Степень сходства и различия между разными зоогео-графическимн областями неодинакова. Фауна и флора Палеоарк-тической и Неоарктической областей имеют много общего, хотя и изолированы Беринговым проливом- Неоарктическая и Неотропическая области, наоборот, существенно отличаются, хотя и соединены сухопутной связью (Панамским перешейком). В чем же причины этого сходства и различия? Очевидно, они связаны с историей формирования материков, временем их изоляции. Так, глубокое различие в фауне Неотропической и Неоарктической областей определяется тем, что сухопутная связь между ними установилась совсем недавно. Об этом свидетельствуют геологические данные.
После возникновения Панамского моста лишь немногим южноамериканским видам удалось проникнуть на север (например, дикобраз, броненосец, опоссум). Североамериканские виды преуспели в освоении южноамериканской области несколько больше. Олени, лисы, выдры, медведи проникли в Южную Америку, но не оказали существенного влияния на ее уникальный видовой состав. Только здесь живут представители отряда неполнозубых (муравьеды и ленивцы), сохранились птицы гоацины, которые могут лазать по деревьям благодаря когтям на пальце крыла (как археоптерикс), и другие животные. Своеобразие фауны Южной Америки было одним из фактов, натолкнувших Ч. Дарвина на мысль об эволюции. Сходство фауны Неоарктической и Палеоарктической областей обусловлено тем, что в прошлом между ними существовал сухопутный мост — Берингов перешеек.
Наиболее отличен от других континентов животный мир Австралии. Известно, что Австралия обособилась от Южной Азии свыше 100 млн. лет назад, еще до возникновения высших млекопитающих. Лишь в Ледниковый период сюда через острова Зонд-ского архипелага перебрались немногие плацентарные — мыши и собаки (потомок последних—австралийская собака динго). В остальном фауна Австралии очень своеобразна.
Таким образом, чем теснее связь континентов, тем более родственные формы там обитают, чем древнее изоляция частей света друг от друга, тем больше различия между их
населением.
Фауна островов
Видовой состав фауны и флоры островов целиком определяется историей их происхождения. Острова могут быть материковыми, представлять собой результат обособления части материка и океаническими (вулканические и коралловые острова). Растительный и животный мир первых близок по составу к материковому. На Британских островах и на Сахалине большинство видов сходно с близлежащими районами континента. Это объясняется тем, что острова лишь несколько тысяч лет назад отделились от суши. Однако чем древнее остров и чем более значительна водная преграда, тем больше обнаруживается отличий. На Мадагаскаре нет типичных для Африки крупных копытных:
быков, антилоп, зебр, нет и крупных хищников (львов, леопардов, гиен), высших обезьян. Однако Мадагаскар — последнее убежище лемуров. Когда-то, до появления обезьян, лемуры были доминирующими приматами. Но они не могли соперничать со своими более развитыми сородичами и исчезли повсюду, кроме Мадагаскара, который отделился от материка прежде, чем эволюционировали обезьяны. Другая большая группа мадагаскарских животных — тенреки и виверры — древние, мало изменившиеся за длительный период изоляции примитивные плацентарные млекопитающие. Мадагаскар обладает широким разнообразием мест обитания, и неудивительно, что он приютил большое количество различных птиц, 46 родов которых не встречаются больше нигде в мире. Трудно представить себе более причудливое животное, чем мадагаскарский хамелеон. Хамелеоны обитают и в Африке, но мадагаскарские виды крупнее и разнообразнее. К ним принадлежит самый большой хамелеон Устале длиной 60 см. У некоторых хамелеонов Мадагаскара на конце морды расположены рога, что делает их похожими на грозных миниатюрных динозавров. Любопытно, что при изобилии ядовитых змей на африканском континенте они отсутствуют на острове. Здесь широко представлены питоны и другие неядовитые змеи. Согласно истории живого мира змеи появились довольно поздно по сравнению с другими рептилиями, причем ядовитые змеи — самые из них молодые. Не значит ли это, что Мадагаскар отделился от континента до появления там змей? Мадагаскар — прекрасное место для лягушек, которых здесь насчитывается около 150 видов.
Иная картина обнаруживается при рассмотрении фауны океанических островов. Ее видовой состав беден и является результатом случайного занесения некоторых видов, обычно птиц, рептилий, насекомых. Наземные млекопитающие, амфибии и другие животные, не способные преодолевать значительные водные преграды, на большинстве таких островов отсутствуют. Приведем пример. Галапагосские острова удалены от берегов Южной Америки на 700 км. Это расстояние могут преодолеть только хорошо летающие формы. Оказалось, что 15% видов птиц Галапагоса представлены южноамериканскими видами, а 85% видов птиц отличны от материковых и нигде, кроме этого архипелага, не встречаются. Именно это с удивлением и наблюдал Ч. Дарвин на Галапагосских островах. Он обнаружил там птиц, которые хотя и напоминали вьюрков, виденных им в Южной Америке, но все были специализированы и мало похожи друг на друга.
У этих 14 видов птиц, имевших общего предка, конкурентная борьба за корм и места гнездования была ослаблена или исчезла вовсе. Ч. Дарвин провел лишь три недели на раскаленных скальных островах. однако увиденное послужило ему материалом для размышлений на долгие годы. Особенно пристальное внимание он обратил на клюв вьюрков: у одних клюв был толстый, дробящий, у других — длинный, у третьих — острый, как ножницы. К одному из шести родов относится дятловый древесный вьюрок, одно ия редких в мире животных, использующих орудия. К сожалению, Ч. Дарвин не увидел эту замечательную птицу заработой, когда она колючкой кактуса выковыривает из древесной коры насекомых и личинок, подобно тому, как это делают дятлы клювом. Галапагосские вьюрки вошли в историю как первый пример влияния изоляции на развитие видов. Сегодня таких примеров известно много. Ч. Дарвин не побывал на Гавайских островах, где эффект изоляции проявляется еще более наглядно на гавайских цветочницах и улитках, прежде всего потому, что они обитают на этих отдаленных островах много дольше, чем вьюрки на Галапагосах.
Дикие лошади
Сегодня настоящих диких лошадей практически не осталось. Российский тарпан считается вымершим видом, а лошадь Пржевальского в естественных условиях не встречается. Табуны лошадей, которые пасутся вольно, — это одичавшие одомашненные формы. Такие лошади хоть и быстро возвращаются к дикой жизни, но все же отличаются от истинно диких лошадей.
У них длинные гривы и хвосты, и они разномастные. Исключительно буланые лошади Пржевальского и мышастые тарпаны имеют как бы подстриженные челки, гривы и хвосты.
В Центральной и Северной Америке дикие лошади были полностью истреблены индейцами и появились там только после прибытия европейцев в XV веке. Одичавшие потомки лошадей конкистадоров дали начало многочисленным табунам мустангов, численность которых сегодня контролируется отстрелом.
Кроме мустангов в Северной Америке есть два вида диких островных пони – на островах Ассатиг и Сейбл. Полудикие табуны лошадей камаргу встречаются на юге Франции. В горах и болотах Британии тоже можно встретить некоторых диких пони.



